Produção melhorada de plasmídeos: Obtenção de um aumento de 21 vezes nos plasmídeos

Introdução
A procura de ADN plasmídico (ADNp) tem aumentado nos últimos anos devido à elevada procura de terapias genéticas e vacinas de ADN (o ADNp é normalmente utilizado devido à sua elevada segurança). Por conseguinte, é altamente necessária uma maior produção de pDNA com um sistema de purificação e controlo de qualidade rentável, reprodutível e fiável. Os plasmídeos são normalmente produzidos em células de Escherichia coli (E. coli) e subsequentemente isolados através de uma série de etapas de purificação. Embora a E. coli produza principalmente a isoforma mais compacta de ADN plasmídico (ADNp) superenrolado (SC), também estão normalmente presentes isoformas de ADNp circular aberto (OC), cortado, linear e desnaturado. A ocorrência de diferentes isoformas pode ser causada por alterações conformacionais que ocorrem no hospedeiro bacteriano e durante o processamento da biomassa (por exemplo, lise celular) e as etapas de purificação do plasmídeo [1]. Várias linhas de evidência indicam que são necessários níveis elevados de SC para desencadear uma resposta imunitária eficaz e, em última análise, para a proteção contra infecções [2,3]. Além disso, a isoforma SC do pDNA é a isoforma desejada para a transfecção, uma vez que permite uma maior eficiência de transfecção devido ao seu empacotamento mais compacto em comparação com as variantes OC ou linear [4-6].
A cromatografia de permuta aniónica (AEC) é um método comum para purificar o SC pDNA de outras isoformas de plasmídeo e para remover as impurezas presentes derivadas do organismo hospedeiro. Idealmente, o processo de produção a montante já fornece predominantemente pDNA SC de alta qualidade.
Materiais e métodos
Transformação e cultivo
O plasmídeo pCMV3-GFP (6883 pb) contendo um gene de resistência à ampicilina (Sino Biological) foi transformado em células E. coli competentes NEB 5-alfa (New England Biolabs) de acordo com o protocolo do fabricante. A mistura de transformação foi colocada numa placa de seleção (LB agar com 100 µg/mL de ampicilina) e incubada durante a noite a 37°C. Escolheu-se uma única colónia da placa de seleção e preparou-se uma cultura de um dia para o outro num Erlenmeyer de vidro de 500 ml, não esvaziado, contendo 50 ml de meio LB (triptona 10 g/L, extrato de levedura 5 g/L, NaCl 10 g/L, pH 7,0 ±0,2) com 100 µg/mL de ampicilina. O frasco foi selado com folha de alumínio e incubado a 37°C e 180 min -1 num agitador de incubadora orbital com tapetes adesivos Sticky Stuff (INFORS HT Multitron Incubator Shaker, 25 mm shaking throw) durante a noite (pelo menos 18 horas). Para a cultura principal, foram preparadas triplicatas de frascos Erlenmeyer de vidro de 250 mL, sem balão, contendo 25 mL de LB com 100 µg/mL de ampicilina, o que é considerado um modelo de cultura padrão.
Foram preparadas triplicatas de frascos Ultra Yield® de 250 ml (Thomson, N/P: 931144) contendo 100 ml de meio Plasmid+® (Thomson, N/P: 446300) com 100 µg/mL de ampicilina e 0,02% de Antifoam 204 (Sigma-Aldrich). Tanto o sistema de cultivo padrão como o sistema de cultivo Thomson foram inoculados a partir da cultura nocturna, com uma OD600nm inicial de 0,2. Os frascos Ultra Yield® foram selados com o AirOtop® Enhanced Seal (Thomson, P/N: 899423) e os frascos Erlenmeyer com folha de alumínio. Os frascos foram incubados a 37°C a 350 min -1 ou 180 min -1 no agitador INFORS HT Multitron Incubator Shaker (25 mm de agitação) com pinças para frascos. Note-se que os tapetes adesivos Sticky Stuff não são adequados para 350 min -1 e a utilização de pinças é obrigatória. As culturas foram cultivadas durante pelo menos 24 horas e foram colhidas amostras a intervalos regulares para medir a OD600nm ao longo do tempo.
Purificação do ADN
O PureYield™ Plasmid Miniprep System (Promega) foi utilizado para a purificação de plasmídeos. A suspensão de células LB foi utilizada sem diluição, as suspensões de células Thomson foram diluídas 9 vezes para corresponder aproximadamente ao valor de DO das amostras LB. Foram colhidas amostras de 600 µL de cada suspensão celular, que foram depois centrifugadas, e o sedimento foi ressuspendido em 600 µL de água sem nuclease. Todos os outros passos subsequentes foram efectuados de acordo com o protocolo rápido do PureYield™ Plasmid Miniprep System FB093 para o kit A1223 ou A1222. A purificação do ADN foi efectuada pelo método de centrifugação. Os plasmídeos foram eluídos com água sem nuclease fornecida com o kit. A concentração e a pureza do ADN das amostras eluídas foram medidas a 230, 260 e 280 nm com um espetrofotómetro NanoDrop™ 2000/2000c (Thermo Fisher Scientific).
Análise de plasmídeos por HPLC
Para a análise de plasmídeos, foi utilizado um HPLC série 1100 (Agilent) com detetor DAD série 1100 (Agilent, G1315B) a 260 nm. As amostras foram medidas numa coluna BioPro IEX QF, 100 x 4,6 mm, 5 µm de tamanho de partícula (YMC, QF00S05-1046WP). Foram injectados 5 µL de cada amostra e medidos em triplicado. A temperatura da coluna foi fixada em 35°C. A fase móvel A (20 mM Tris-HCl, pH 7,4) e a fase móvel B (20 mM Tris-HCl, 1 M NaCl, pH 7,4) foram utilizadas para eluir a amostra num método de gradiente com um fluxo constante de 0,5 mL/min. O gradiente foi efectuado do minuto 0 até ao minuto 0,7 com a fase móvel A (MPA) a 25% e a fase móvel B (MPB) a 75%. A partir do minuto 0.7 o gradiente foi aplicado e a composição de 75% MPB foi alterada para 100% durante 20 minutos até ao minuto 20.7 e foi mantida a 100% até ao minuto 24. No minuto 24.01, a composição foi alterada para 25% de MPA e 75% de MPB até ao minuto 30 (fim da injeção).
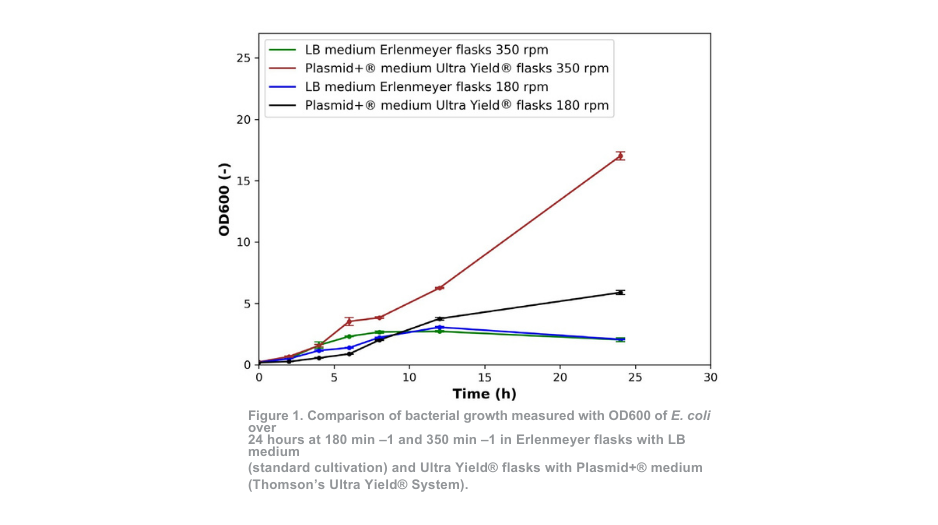
O crescimento bacteriano em frascos Erlenmeyer com meio LB (cultivo padrão) a 350 min-1 e em frascos Ultra Yield® com meio Plasmid+® a 350 min -1 apresentou padrões semelhantes até 4 horas de cultivo, como mostra a Figura 1. Para o cultivo a 180 min -1, as semelhanças são observadas até 8 horas. No caso do crescimento bacteriano em frascos Ultra Yield®, a taxa de crescimento inicial é ligeiramente mais lenta durante as primeiras horas em comparação com a cultura padrão.
No entanto, torna-se evidente um contraste distinto entre as duas condições de cultivo após 4 horas (350 min -1) e 8 horas (180 min -1). Para a abordagem de cultivo padrão, a densidade ótica é reduzida em comparação com a medição às 12 horas, indicando que o número de células viáveis já está a diminuir neste momento. No final do período de cultivo, os valores de DO do sistema Ultra Yield® mantidos a 180 min-1 e 350 min-1 excederam os do sistema de cultivo padrão em mais de 2 vezes e 7 vezes, respetivamente.
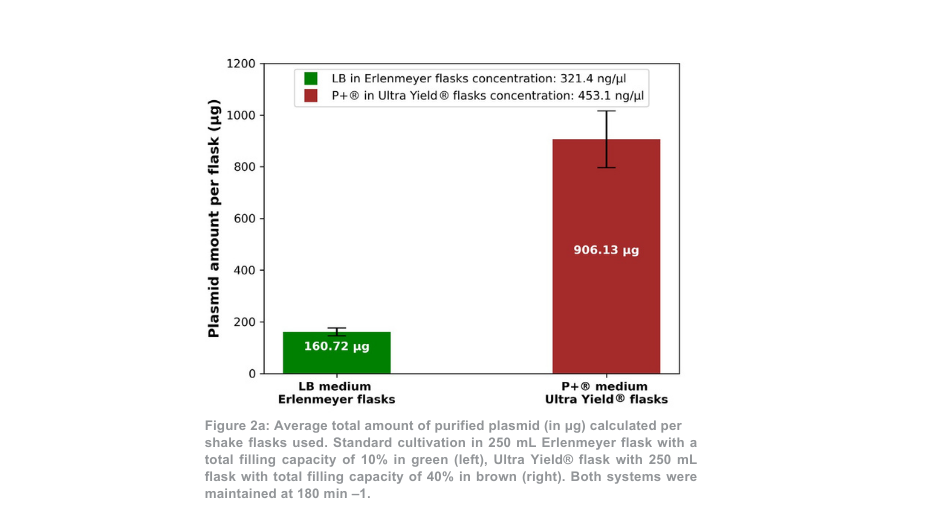
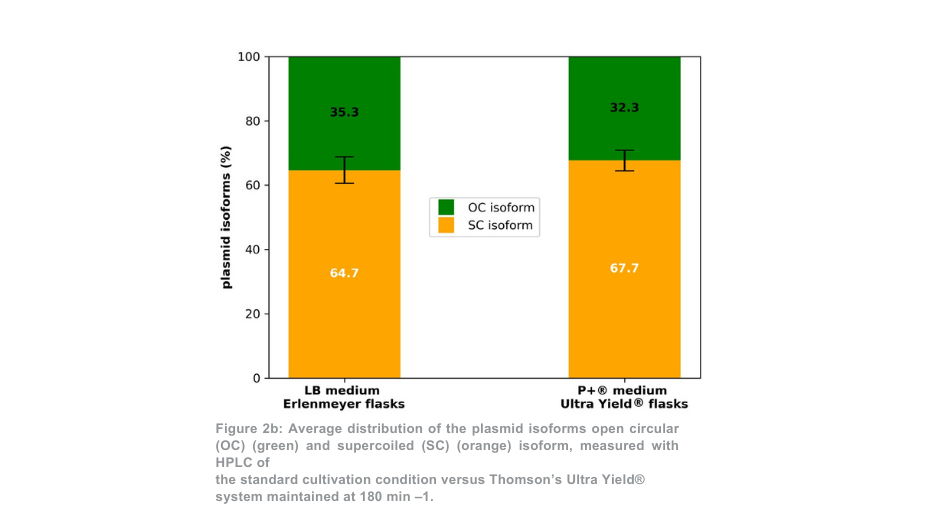
Após a purificação do plasmídeo, a produção total de plasmídeo para a condição de 180 min -1 foi mais de 5 vezes superior por frasco Thomson em comparação com os frascos Erlenmeyer (Figura 2a). Quando a diferença de volume entre os frascos foi contabilizada, a produção de plasmídeo foi 1,4 vezes maior com o sistema de cultivo Thomson em comparação com o sistema de cultivo padrão. A proporção de plasmídeo superenrolado em relação ao plasmídeo enrolado aberto (Figura 2b) foi semelhante entre o sistema de cultivo Thomson (67,7% superenrolado) e o sistema de cultivo padrão (64,7% superenrolado).
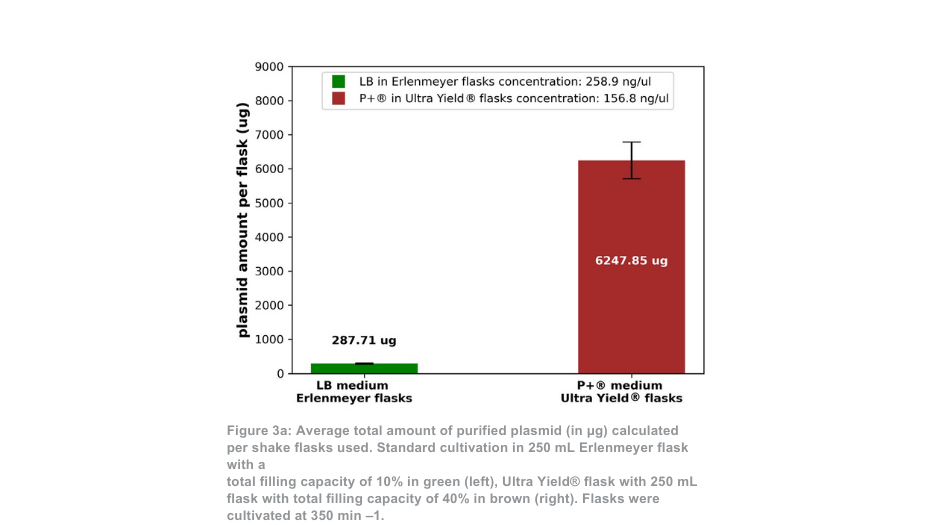
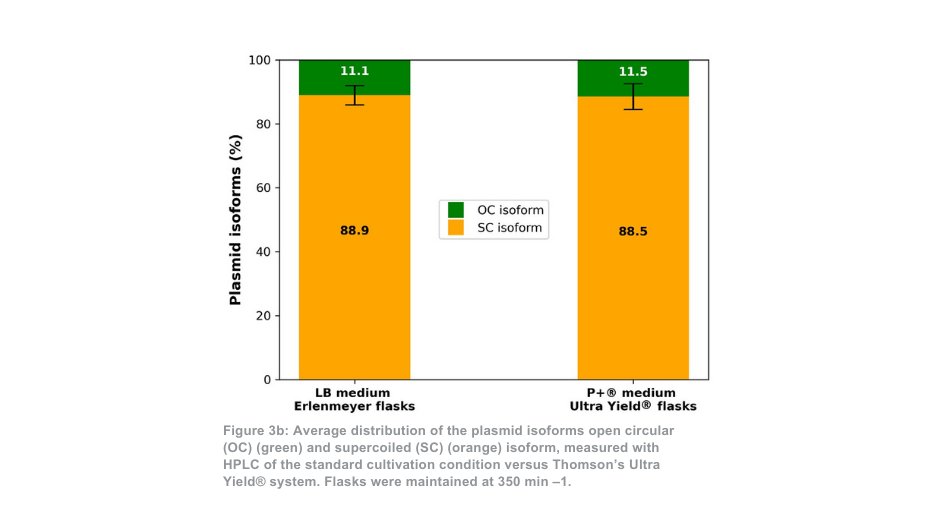
A diferença entre o sistema de cultivo Thomson e o sistema de cultivo padrão é mais pronunciada quando mantido a 350 min -1. Neste caso, a quantidade total de plasmídeo purificado por frasco é cerca de 21 vezes superior com o sistema Ultra Yield® da Thomson em comparação com a condição de cultivo padrão, ou cerca de 5 vezes superior quando se tem em conta a diferença de volume (Figura 3a). A proporção de plasmídeo superenrolado em relação ao plasmídeo enrolado aberto (Figura 3b) foi novamente semelhante entre o sistema de cultivo Thomson (88,5% superenrolado) e o sistema de cultivo padrão (88,9% superenrolado).
Conclusão
Os dados demonstram que as velocidades de agitação comuns de 180 min -1 (órbita de 25 mm) não são suficientes para suportar um crescimento celular ótimo nos frascos Thomson Ultra Yield®. Uma velocidade de agitação elevada de 350 min -1 proporciona um aumento de 3 vezes no rendimento da biomassa, em comparação com 180 min -1. Com um melhor crescimento celular, foi alcançada uma maior produção de pDNA. O sucesso desta aplicação depende em grande medida da manutenção de uma velocidade de agitação elevada para alcançar uma mistura e transferência de gás eficazes, conduzindo a rendimentos mais elevados. À medida que as velocidades de agitação aumentam de 180 min -1 para 350 min -1, há um aumento de 4 vezes na energia cinética necessária para agitar eficazmente a superfície. É fundamental que o limite de carga do agitador possa acomodar com segurança os frascos a altas velocidades. O agitador de incubadora INFORS HT Multitron pode conter 45 frascos de 250 mL, com uma massa de meios e cultura totalizando 4,5 kg. Considerando a massa adicional da bandeja e das pinças dos frascos, o limite mínimo de carga do agitador deve exceder 12 kg. Com frascos maiores, é possível carregar mais de 10 L de meios no agitador, tornando o limite de carga física e as dimensões interiores do agitador extremamente importantes. Todos estes factores enfatizam a necessidade de um agitador robusto, como o agitador de incubadora Multitron, para obter todos os benefícios deste processo.
A combinação dos frascos Ultra Yield® com o meio Plasmid+® enriquecido e o vedante melhorado AirOtop® conduziu a densidades celulares mais elevadas e a uma quantidade de plasmídeos 21 vezes superior à dos frascos Erlenmeyer com meio LB. O rendimento global mais elevado de plasmídeos também resulta numa maior quantidade da isoforma de plasmídeo superenrolada (SC), que é desejada para a transfecção de células de mamíferos [4,5]. Considerando o custo total de propriedade de um processo de produção de plasmídeos no laboratório, a utilização de frascos Thomson Ultra Yield®, meio Plasmid+® em combinação com um potente agitador de incubadora a 350 min -1 é menos dispendiosa em mg/L de plasmídeo em comparação com a cultura LB em Erlenmeyers de vidro. É vantajoso investir num bom agitador para atingir velocidades mais elevadas e no sistema Thomson.
Utilizar AIROTOP® Seal/Vented Screw Cap Media mL/Flask Velocidade do agitador (RPM)
| Código | Descrição | Utilizar a tampa de rosca com vedação/ventilação AIROTOP® | Meio mL/Frasco | Velocidade do agitador (RPM) |
| 931147 | Frascos Ultra Yield® de 125 ml 50/CS - Estéreis | 899421 / 899109 | 35-50 mL/Flask | 300-350 |
| 931144 | Frascos Ultra Yield® de 250 ml 50/CS - Estéreis | 899423 / 899110 | 75-100 mL/Flask | 300-350 |
| 931141 | Frascos Ultra Yield® de 500 ml 25/CS - Estéreis | 899424 / 899111 | 150-200 mL/Flask | 300-350 |
| 931138 | Garrafas de 1,5L Ultra Yield® 12/CS - Estéreis | 899425 / 899566 | 300 mL/Flask | 300-350 |
| 931136-B | Garrafas Ultra Yield® de 2,5 L 6/CS - Estéreis | 899425 / 899566 | 500 mL/Flask | 300-400 |
| 446300 | Plasmid+® Enriched Media - Estéril | N/A | N/A | N/A |
Referências
- D.M. Prazeres, T. Schluep, C. Cooney, Purificação preparativa de ADN plasmídico superenrolado por cromatografia de permuta aniónica, J. Chromatogr. A 806 (1998) 31–45. https://doi.org/10.1016/s0021-9673(97)01254-5.
- Lionel Cupillard, Véronique Juillard, Sophie Latour, Guy Colombet, N. Cachet, S. Richard, S. Blanchard, Laurent Fischer, Impacto do enrolamento do plasmídeo na eficácia de uma vacina de ADN contra a raiva para proteger os gatos, Vaccine 23 (2005) 1910–1916. https://doi.org/10.1016/J.VACCINE.2004.10.018.
- 3. Huangjin Li, Huaben Bo, Jinquan Wang, Hongwei Shao, Shulin Huang, Separação de formas circulares superenroladas e abertas de ADN plasmídico e deteção de atividade biológica, Cytotechnology 63 (2011) 7–12. https://doi. org/10.1007/s10616-010-9322.
- Christof Maucksch, Alexander Bohla, Florian Hoffmann, Martin Schleef, Manish Kumar Aneja, Markus Elfinger, Dominik Hartl, Carsten Rudolph, Expressão transgénica de concâmeros de ADN plasmídico superenrolado transfectados em células de mamíferos, A revista de medicina genética 11 (2009) 444–453. https://doi.org/10.1002/jgm.1310.
- Arjun Dhanoya, Benjamin M. Chain, Eli Keshavarz-Moore, O impacto da topologia do ADN na captação de poliplexos e na eficiência da transfecção em células de mamíferos, Jornal de Biotecnologia 155 (2011) 377–386. https://doi.org/10.1016/J.JBIOTEC.2011.07.023.
- Fani Sousa, Duarte M.F. Prazeres, João A. Queiroz, Melhoria da eficiência da transfecção através da utilização de ADN plasmídico superenrolado purificado por cromatografia de afinidade com arginina, A revista de medicina genética 11 (2009) 79–88. https://doi.org/10.1002/JGM.1272.