Producción mejorada de plásmidos: Logrando un Aumento de 21 Veces en Plásmidos

Introducción
La demanda de ADN plasmídico (ADNp) ha aumentado en los últimos años como consecuencia de la gran demanda de terapias génicas y vacunas de ADN (el ADNp se utiliza habitualmente por su gran seguridad). Por lo tanto, es muy necesaria una mayor producción de ADNp con un sistema de purificación y control de calidad rentable, reproducible y fiable. Normalmente, los plásmidos se producen en células de Escherichia coli (E. coli) y posteriormente se aíslan mediante una serie de pasos de purificación. Aunque E. coli produce principalmente la isoforma más compacta de ADN plasmídico (ADNp) superenrollado (SC), también suelen estar presentes las isoformas de ADNp circular abierto (OC), mellado, lineal y desnaturalizado. La aparición de diferentes isoformas puede deberse a cambios conformacionales que se producen en el huésped bacteriano y durante el procesamiento de la biomasa (por ejemplo, la lisis celular) y los pasos de purificación del plásmido [1]. Varias líneas de evidencia indican que se requieren altos niveles de SC para provocar una respuesta inmunitaria eficaz y, en última instancia, protección frente a las infecciones [2,3]. Además, la isoforma SC pDNA es la isoforma deseada para la transfección, ya que permite una mayor eficiencia de transfección debido a su empaquetamiento más compacto en comparación con las variantes OC o lineales [4-6].
La cromatografía de intercambio aniónico (CEA) es un método habitual para purificar el ADNp de SC de otras isoformas plasmídicas y para eliminar las impurezas presentes derivadas del organismo huésped. Lo ideal es que el proceso de producción previo ya proporcione pADN de SC predominantemente de alta calidad.
Materiales y métodos
Transformación y cultivo
El plásmido pCMV3-GFP (6883 pb) que contiene un gen de resistencia a la ampicilina (Sino Biological) se transformó en células E. coli NEB 5-alfa competentes (New England Biolabs) siguiendo el protocolo del fabricante. La mezcla de transformación se sembró en una placa de selección (agar LB con 100 µg/mL de ampicilina) y se incubó toda la noche a 37°C. Se escogió una sola colonia de la placa de selección y se preparó un cultivo de una noche en un matraz Erlenmeyer de vidrio de 500 mL sin esponjar que contenía 50 mL de medio LB (triptona 10 g/L, extracto de levadura 5 g/L, NaCl 10 g/L, pH 7,0 ±0,2) con 100 µg/mL de ampicilina. El matraz se selló con papel de aluminio y se incubó a 37°C y 180 min -1 en un agitador de incubadora orbital con esterillas adhesivas Sticky Stuff (Agitador de incubadora INFORS HT Multitron, tiro de agitación de 25 mm) durante toda la noche (al menos 18 horas). Para el cultivo principal, se prepararon triplicados de matraces Erlenmeyer de vidrio de 250 mL sin esponjar que contenían 25 mL de LB con 100 µg/mL de ampicilina, lo que se considera un diseño de cultivo estándar.
Se prepararon triplicados de matraces Ultra Yield® de 250 mL (Thomson, N/P: 931144) que contenían 100 mL de medio Plasmid+® (Thomson, N/P: 446300) con 100 µg/mL de ampicilina y 0,02% de Antifoam 204 (Sigma-Aldrich). Tanto el sistema de cultivo estándar como el sistema de cultivo Thomson se inocularon a partir del cultivo de una noche, con una DO600nm inicial de 0,2. Los matraces Ultra Yield® se sellaron con el AirOtop® Enhanced Seal (Thomson, P/N: 899423) y los matraces Erlenmeyer con papel de aluminio. Los matraces se incubaron a 37°C a 350 min -1 o 180 min -1 en el Agitador Incubador Multitron HT de INFORS (tiro de agitación de 25 mm) con pinzas para matraces. Tenga en cuenta que las esteras adhesivas Sticky Stuff no son adecuadas para 350 min -1 y que es obligatorio el uso de pinzas. Los cultivos se cultivaron durante al menos 24 horas y se tomaron muestras a intervalos regulares para medir la OD600nm a lo largo del tiempo.
Purificación del ADN
Para la purificación de plásmidos se utilizó el sistema PureYield™ Plasmid Miniprep (Promega). La suspensión celular de LB se utilizó sin diluir, las suspensiones celulares de Thomson se diluyeron 9 veces para igualar aproximadamente el valor DO de las muestras de LB. Se tomaron muestras de 600 µl de cada suspensión celular, que luego se centrifugaron y el sedimento se resuspendió en 600 µl de agua libre de nucleasas. Todos los demás pasos posteriores se llevaron a cabo de acuerdo con el protocolo rápido FB093 del sistema de minipreparación de plásmidos PureYield™ para el kit A1223 o A1222. La purificación del ADN se realizó por el método de centrifugación. Los plásmidos se eluyeron con agua libre de nucleasas suministrada con el kit. La concentración de ADN y la pureza de las muestras eluidas se midieron a 230, 260 y 280 nm con un espectrofotómetro NanoDrop™ 2000/2000c (Thermo Fisher Scientific).
Análisis de plásmidos mediante HPLC
Para el análisis de plásmidos se utilizó un HPLC Serie 1100 (Agilent) con detector DAD Serie 1100 (Agilent, G1315B) a 260 nm. Las muestras se midieron en una columna BioPro IEX QF, 100 x 4,6 mm, 5 µm de tamaño de partícula (YMC, QF00S05-1046WP). Se inyectaron 5 µl de cada muestra y se midieron por triplicado. La temperatura de la columna se fijó en 35°C. Se utilizó la fase móvil A (20 mM Tris-HCl, pH 7,4) y la fase móvil B (20 mM Tris-HCl, 1 M NaCl, pH 7,4) para eluir la muestra en un método de gradiente con un flujo constante de 0,5 mL/min. El gradiente se realizó desde el minuto 0 hasta el minuto 0,7 con la fase móvil A (MPA) al 25% y la fase móvil B (MPB) al 75%. A partir del minuto 0,7 se aplicó el gradiente y la composición de MPB al 75% se cambió al 100% durante 20 minutos hasta el minuto 20,7 y se mantuvo al 100% hasta el minuto 24. En el minuto 24,01 la composición se cambió al 100%. En el minuto 24,01 se cambió la composición a 25% de MPA y 75% de MPB hasta el minuto 30 (fin de la inyección).
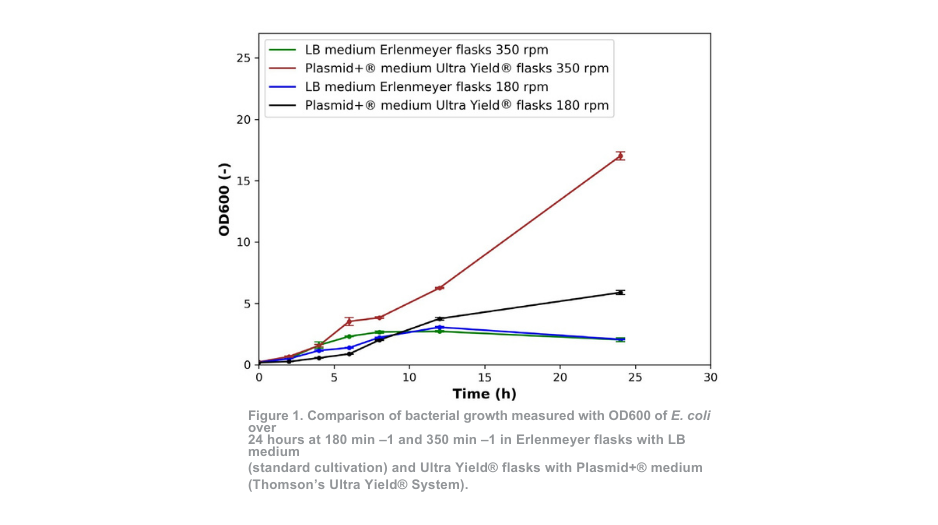
El crecimiento bacteriano en matraces Erlenmeyer con medio LB (cultivo estándar) a 350 min-1 y en matraces Ultra Yield® con medio Plasmid+® a 350 min -1 mostró patrones similares hasta las 4 horas de cultivo, como se muestra en la figura 1. Para el cultivo a 180 min -1 las similitudes se observan hasta las 8 horas. En el caso del crecimiento bacteriano en matraces Ultra Yield®, la tasa de crecimiento inicial es ligeramente más lenta durante las primeras horas en comparación con el cultivo estándar.
Sin embargo, se hace evidente un claro contraste entre las dos condiciones de cultivo después de 4 horas (350 min -1) y 8 horas (180 min -1). En el método de cultivo estándar, la densidad óptica se reduce en comparación con la medición realizada a las 12 horas, lo que indica que el número de células viables ya está disminuyendo en ese momento. Al final del periodo de cultivo, los valores de DO del sistema Ultra Yield® mantenido a 180 min-1 y 350 min-1 superaron a los del sistema de cultivo estándar en más de 2 veces y 7 veces respectivamente.

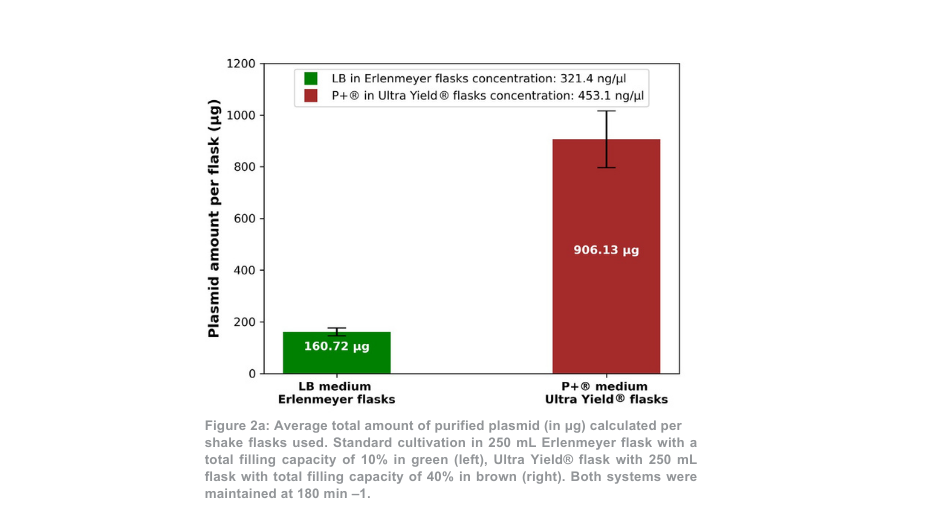
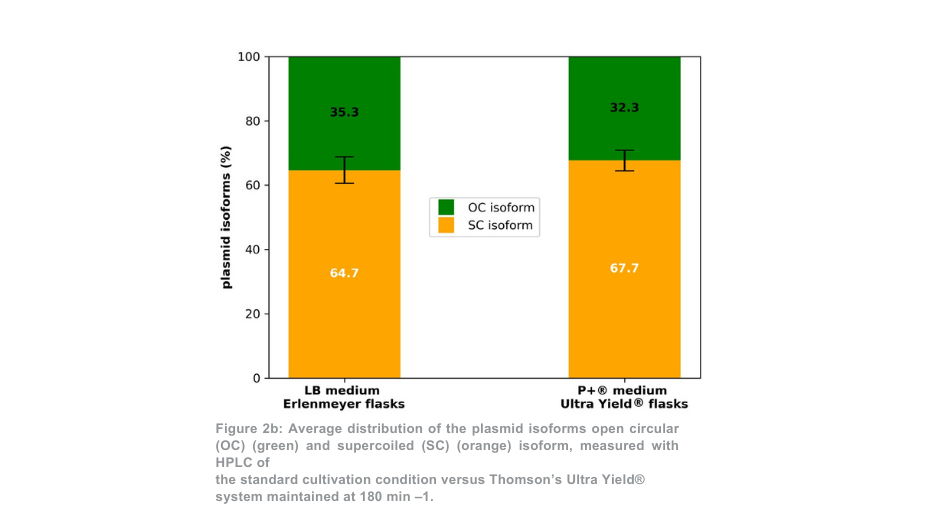
Tras la purificación de plásmidos, el rendimiento total de plásmidos para la condición de 180 min -1 fue más de 5 veces superior por matraz Thomson en comparación con los matraces Erlenmeyer (Figura 2a). Cuando se tuvo en cuenta la diferencia de volumen entre los matraces, el rendimiento de plásmidos fue 1,4 veces mayor con el sistema de cultivo Thomson que con el sistema de cultivo estándar. La proporción de plásmido superenrollado respecto al plásmido en espiral abierto (Figura 2b) fue similar entre el sistema de cultivo Thomson (67,7% superenrollado) y el sistema de cultivo estándar (64,7% superenrollado).
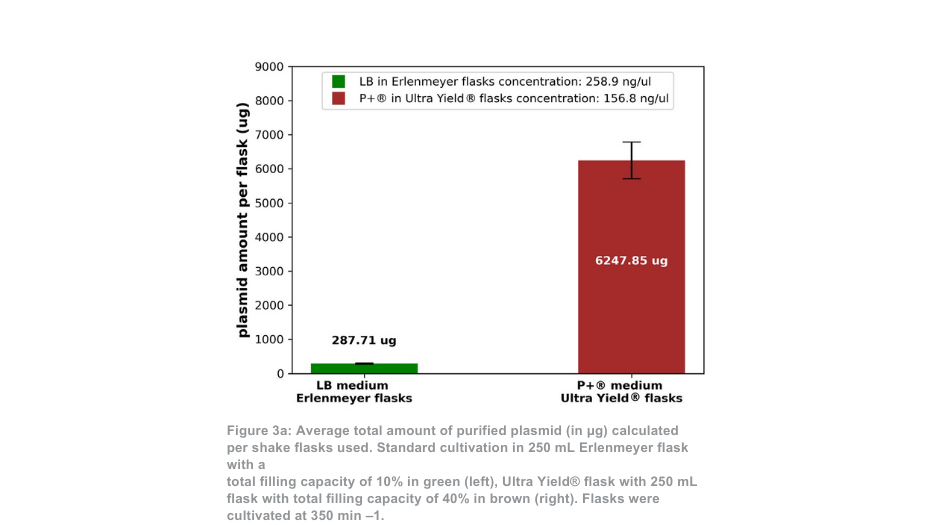
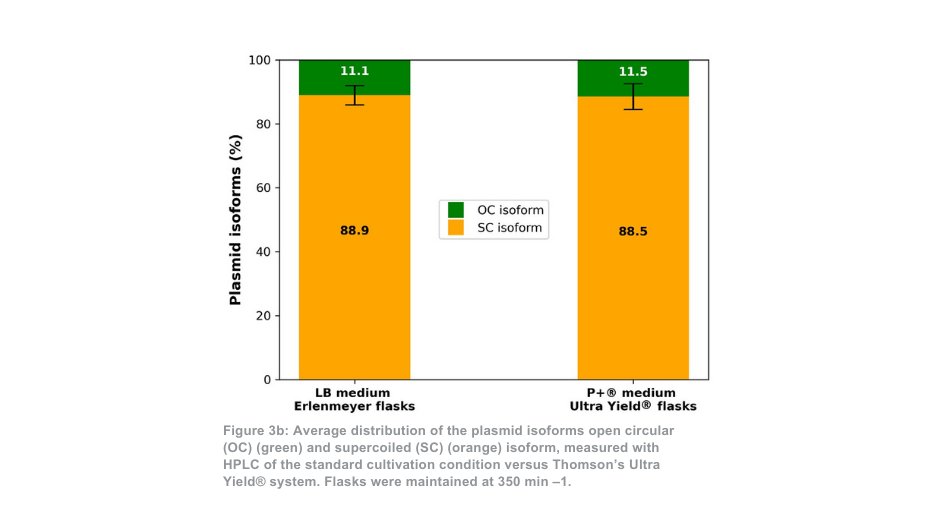
La diferencia entre el sistema de cultivo Thomson y el sistema de cultivo estándar es más pronunciada cuando se mantiene a 350 min -1 . En este caso, la cantidad total de plásmido purificado por matraz es aproximadamente 21 veces mayor con el sistema Ultra Yield® de Thomson en comparación con la condición de cultivo estándar, o aproximadamente 5 veces mayor si se tiene en cuenta la diferencia de volumen (Figura 3a). La proporción de plásmido superenrollado con respecto al plásmido en espiral abierto (Figura 3b) fue de nuevo similar entre el sistema de cultivo de Thomson (88,5% superenrollado) y el sistema de cultivo estándar (88,9% superenrollado).
Conclusión
Los datos demuestran que las velocidades de agitación habituales de 180 min -1 (órbita de 25 mm) no son suficientes para mantener un crecimiento celular óptimo en los matraces Ultra Yield® de Thomson. Una velocidad de agitación elevada de 350 min -1 multiplica por 3 el rendimiento de la biomasa, en comparación con 180 min -1. Con un mejor crecimiento celular, la producción de ADNp es mayor. Con un mejor crecimiento celular, se consiguió una mayor producción de ADNp. El éxito de esta aplicación depende en gran medida de mantener una velocidad de agitación elevada para lograr una mezcla y una transferencia de gases eficaces, lo que conduce a mayores rendimientos. A medida que aumenta la velocidad de agitación de 180 min -1 a 350 min -1, se multiplica por 4 la energía cinética necesaria para agitar eficazmente la superficie. Es fundamental que el límite de carga del agitador pueda acomodar con seguridad los matraces a altas velocidades. El agitador incubador Multitron HT de INFORS tiene capacidad para 45 matraces de 250 ml, con una masa de medios y cultivos que asciende a un total de 4,5 kilogramos. Teniendo en cuenta la masa adicional de la bandeja y las abrazaderas de los matraces, el límite mínimo de carga del agitador debería superar los 12 kilogramos. Con matraces más grandes, es posible cargar más de 10 L de medio en el agitador, por lo que el límite de carga física y las dimensiones interiores del agitador son extremadamente importantes. Todos estos factores enfatizan la necesidad de un agitador robusto, como el Multitron Incubator Shaker, para aprovechar plenamente los beneficios de este proceso.
La combinación de los matraces Ultra Yield® con el medio enriquecido Plasmid+® y el cierre mejorado AirOtop® permitió obtener densidades celulares más altas y una cantidad de plásmidos 21 veces mayor que en los matraces Erlenmeyer con medio LB. El mayor rendimiento global de plásmidos también se traduce en una mayor cantidad de la isoforma superenrollada (SC) del plásmido, que es la deseada para la transfección de células de mamífero [4,5]. Teniendo en cuenta el coste total de propiedad de un proceso de producción de plásmidos en el laboratorio, el uso de los matraces Ultra Yield® de Thomson, el medio Plasmid+® en combinación con un potente agitador incubador a 350 min -1 es menos costoso en mg/L de plásmidos comparado con el cultivo LB en Erlenmeyers de vidrio. Invertir en un buen agitador para alcanzar velocidades más altas, y el sistema Thomson es ventajoso.
Utilice AIROTOP® Seal/Vented Screw Cap Media mL/Flask Velocidad del agitador (RPM)
| Código | Descripción | Utilizar junta AIROTOP®/tapón de rosca ventilado | Medio mL/Frasco | Velocidad del agitador (RPM) |
| 931147 | Matraces Ultra Yield® de 125mL 50/CS - Estéril | 899421 / 899109 | 35-50 mL/Flask | 300-350 |
| 931144 | Matraces Ultra Yield® 250mL 50/CS - Estéril | 899423 / 899110 | 75-100 mL/Flask | 300-350 |
| 931141 | Botellas 500mL Ultra Yield® 25/CS - Estériles | 899424 / 899111 | 150-200 mL/Flask | 300-350 |
| 931138 | Botellas Ultra Yield® 1,5L 12/CS - Estériles | 899425 / 899566 | 300 mL/Flask | 300-350 |
| 931136-B | Botellas 2,5L Ultra Yield® 6/CS - Estériles | 899425 / 899566 | 500 mL/Flask | 300-400 |
| 446300 | Plasmid+® Enriched Media - Estéril | N/A | N/A | N/A |
Referencias
- D.M. Prazeres, T. Schluep, C. Cooney, Purificación preparativa de ADN plasmídico superenrollado mediante cromatografía de intercambio aniónico,J. Chromatogr. A 806 (1998) 31–45. https://doi.org/10.1016/s0021-9673(97)01254-5.
- Lionel Cupillard, Véronique Juillard, Sophie Latour, Guy Colombet, N. Cachet, S. Richard, S. Blanchard, Laurent Fischer, Impacto del superenrollamiento del plásmido en la eficacia de una vacuna de ADN contra la rabia para proteger a los gatos, Vacuna 23 (2005) 1910–1916. https://doi.org/10.1016/J.VACCINE.2004.10.018.
- 3. Huangjin Li, Huaben Bo, Jinquan Wang, Hongwei Shao, Shulin Huang, Separación de las formas circulares superenrolladas de las abiertas del ADN plasmídico y detección de la actividad biológica, Citotecnología 63 (2011) 7–12. https://doi. org/10.1007/s10616-010-9322.
- Christof Maucksch, Alexander Bohla, Florian Hoffmann, Martin Schleef, Manish Kumar Aneja, Markus Elfinger, Dominik Hartl, Carsten Rudolph, Expresión transgénica de concámeros de ADN plasmídico superenrollado transfectados en células de mamífero, La revista de medicina genética 11 (2009) 444–453. https://doi.org/10.1002/jgm.1310.
- Arjun Dhanoya, Benjamin M. Chain, Eli Keshavarz-Moore, Efectos de la topología del ADN en la captación de poliplejos y la eficacia de la transfección en células de mamífero, Revista de Biotecnología 155 (2011) 377–386. https://doi.org/10.1016/J.JBIOTEC.2011.07.023.
- Fani Sousa, Duarte M.F. Prazeres, João A. Queiroz, Mejora de la eficacia de la transfección mediante el uso de ADN plasmídico superenrollado purificado con cromatografía de afinidad por arginina, La revista de medicina genética 11 (2009) 79–88. https://doi.org/10.1002/JGM.1272.